リンパ細胞V(D)J再配列に不可欠な遺伝子―Rag1、Rag2
研究方向を調整し、学術研究の突破口を目指す?科学発展の動向と将来方向性を明確する必要がある!サイヤジェン株式会社のコラム「Gene of the Week」では、毎週火曜日に注目が高い研究分野に基づき1種の遺伝子を紹介し、基礎情報、研究概要と応用を詳しく述べ、研究力と科学研究効率の向上をサポートいたします。そして今回ご紹介するのは、リンパ細胞V(D)J再配列に不可欠な遺伝子―Rag1、Rag2です。
Rag1、Rag2概要
我々はB細胞によって産生される免疫グロブリン(Ig)-B細胞受容体(B cell receptor,BCR)と抗体、およびT細胞上のT細胞表面受容体(T cell receptor, TCR)は共に特異性をもっていることを分かっている;一種のIgまたはTCRタンパク質は特異的に一種の抗原しか識別できない。ただ全体的には、これらのタンパク質は膨大な多様性をもち、この多様性はV(D)J再配列(recombination)によって達成できる。V(D)J再配列は、脊椎動物の初期T細胞とB細胞の発育と成熟の過程での体細胞組換えメカニズムである。そして可変区(variable,V)、多様区(diversity,D)と結合区(joining,J)の遺伝子の組換えを通して、様々な抗原を認識できる多様性IgとTCRが生成される。組換え活性化遺伝子(recombination-activating genes,RAGs):Rag1とRag2は、Rag1とRag2のリコンビナーゼ(recombinases)を其々コードする。Rag1とRag2は、発育中のリンパ細胞の中で制限的に発現されており、適応性免疫システムの重要な部分である。
12/23規則とは
IgとTCRの生殖細胞系遺伝子(germ line gene)のV、D、J区域の両端には共に再配列シグナル配列(recombination signal sequence,RSS)が存在する。構造的にすると、RSSは一端に一つの7bpの配列(heptamer)、もう一端には9bpの配列(nonamer)があり、両者の間に12bpまたは23bpのspacer配列を挟くことから、RSSは2つのタイプに分けることができる:12bpのspacer付き12-RSS、および23bpのspacer付き23-RSS(図1a)である。12/23規則とは、V(D)Jの再配置中に、12-RSSは23-RSSとのみ組み合わせられることから、遺伝子フラグメントの正確な接続を保証できる。例えば、Ig重鎖(IgH)の生殖細胞系遺伝子では、VHおよびJH遺伝子の両側は共に23-RRSであるが、DH遺伝子の両側は12-RSSであり、12/23規則によってVHとJH遺伝子の直接接続を防ぎ、DH遺伝子も再配列に関与されるよう保証できる(図1b)。さらに、分子レベルから考察すると、12-RSSと23-RSSの結合もより安定したシナプス複合体(synapsis)を形成することができる。
V(D)J再配列の過程で、まず、Rag1/2複合体は高移動度群タンパク質B1(high mobility group box 1,HMGB1)の助けを得て(DNA結合を支援)、12-RSSまたは23-RSSを認識し結合し、RSSのheptamerとエンコード端の接合部に一本鎖切開を正確に導入する。切開により、コーディング末端に遊離の3'-OHゲインが生成され、このゲインは逆平行鎖のホスホジエステル結合を攻撃し、ヘアピン構造(hairpin)と鈍性のシグナル末端が形成される。シグナル末端はRAG1 / 2複合体との結合を保ち、ひとつの「post-cleavage complex」の一時的構造を形成する。それから、コーディングエンドとシグナルエンドの双方は共に非同源末端の連結(non-homologous end joining,NHEJ)経路を介して加工と連結を行い、コーディングジョイント(coding joint)と環状シグナルジョイント(signal joint)を形成する(図1c)。
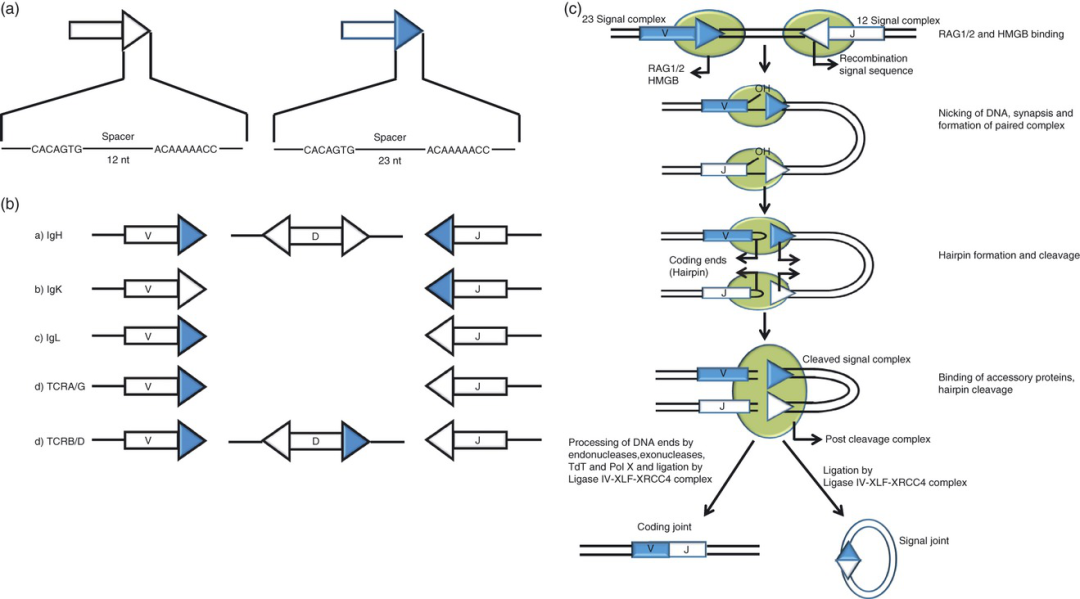
図1.V(D)J再配置
(a)RSS構造:12-RSSと23-RSS:7bpheptamer及び9bpnonamerの間に其々12bpsのpacerまたは23bのpspacerを挟む。
(b)ヒトIgH、IgK、IgL、およびTCRα鎖、β鎖、γ鎖およびδ鎖の遺伝子におけるRSSの分布。
(c)V(D)J再配置メカニズム。
Rag1とRag2の区別
Rag1とRag2の機能上の違いは?Dr.Akamatsuの実験によると、RSSと非特異的DNA配列の存在下では、Rag1とRSSの結合は非特異的DNA配列より僅か3〜5倍高いことが示されている。Rag2に関しては、実験でDNAとの結合を観察できない;ただしRag1とRag2が同時に存在する場合、それらはDNAとRag1-Rag2-DNA複合体を形成できるが、これはRag1-DNA複合体よりも更に安定しており、もっと特異性をもつ。そしてV(D)J組換えにおいて切断活性を持っている。これらの結果は、V(D)J再配列において、Rag2はRag1とDNAを識別する機能を支援する役割を果たしている可能性を示しており、効果的にRSSと結合し認識するにはRag1とRag2の同時存在が必要となる[2]。
まとめ
Rag1とRag2のリコンビナーゼは共にT細胞とB細胞の免疫グロブリンを発育させ、成熟させる重要なタンパク質である。V(D)J再配列では、Rag1/2複合体がRSSの認識とスプライシングに関与している。従って、PrkdcSCID遺伝子変異を持つマウスと比べ、Rag1とRag2のノックアウトマウスはV(D)J再配列経路が最初から阻止されているため,マウスは機能性TとBリンパ細胞が少なく、免疫漏出(“leakiness”)も発生されない。つまり、マウスの体内で少量の機能性TとB細胞および免疫グロブリンが生成される。異なる遺伝背景のSCIDマウスの免疫漏出の発生率は異なり;一般的に、SCIDマウスはC57BL/6JおよびBALB/c背景上での漏出率が高く;C3H背景上では低く;NOD背景上では極めて低い。しかし、現時点では免疫漏出の分子メカニズムはまだ不明である。
参考文献:
[1] Nishana, M., & Raghavan, S. C. (2012). Role of recombination activating genes in the generation of antigen receptor diversity and beyond. Immunology, 137(4), 271–281.
[2] Akamatsu Y, Oettinger MA. (1998). Distinct roles of RAG1 and RAG2 in binding the V(D)J recombination signal sequences. Mol Cell Biol, 18(8):4670-8.
サイヤジェン株式会社について
サイヤジェン株式会社は15年間の発展を経て、全世界の数万人の科学研究者にサービスを提供しており、製品と技術は直接にCNS (Cell、Nature、Science)の定期刊を含む5,200余りの学術論文に応用されています。弊社の「ノックアウトマウスライブラリ」は低価格だけでなく、遺伝子名称を入力すれば、ワンクリックで注文まで操作できます。ノックアウトマウス、ノックインマウス、コンディショナルノックアウトマウス、トランスジェニックマウス、GFPマウス、免疫不全マウス、無菌マウスなどのカスタマイズサービスを提供する以外、専門的な手術疾患モデルチームがあり、多種の複雑な小動物手術疾患モデルも提供できます。
