アルツハイマー病研究の新しい構想——ラットモデルに焦点を絞る
アルツハイマー病(AD)は今の研究用に主な動物モデルの遺伝子改変マウスであり、良い成果を上げたが、相応の薬物実験は多くの失敗がある。特にAβ仮説を中心にする薬物開発はほぼ遭難したとも言える。このような状況で、弊社の構想を生み出し、別のモデルを選別し、ほかの角度からAD薬物開発を見直すべきか。これは数十年間以来ずっとマウスの“影の下”のラットをAD研究の最前線に押した。これは主に以下のいくつかの点である:
1. ラットは全体的にマウスより賢く、飼育しやすく、より複雑な実験を行う可能になる。これまでの複雑な実験遺伝子の改変や薬物に反応しやすくなる。
2. マウスよりラットは人を恐れず、焦慮と陰鬱になりにくくて、神経生物学の行動学実験にもっと適用する。
3. ラットの各組織、器官はマウスより大きく、手術操作も容易になる。
4. AD関連の遺伝子、例えばMAPTがラットでの多型で、より人間に近い。
5. 日増しに容易になるラットの遺伝子改変。
ラットをADの実験動物として、マウスの実験で得られない情報が得られる可能性が高いので、一部の学者はAD研究をラットに選択している。
1. APPのせん断とAβ
長年にわたってADの研究につれて、ADの2つの特徴を見つけた。①患者の脳にAβ(Amyloid Beta)によって集められたデンプンのような沈殿物とtauによって集められた神経線維の絡み合いである。その中で、AβはAPP(Amyloid Precursor Protein)が2回せん断で形成されている:非アミロイドせん断とアミロイドせん断である。前者は毒性Aβを発生しない「良い」せん断方式と考えられ、後者は過剰活発になるとAβが積みすぎてデンプン状沈殿が産生する。この2種類のせん断とも2つの「ハサミ」が必要である。違いは第1目のハサミが異なっている。前者の第1目のカットはα-secretase(αカット酵素)によって行う。αカット酵素はほとんどAPPのAβ断片の中間からせん断するため、Aβ-カット!しかし、注意すべきなのはこのようなせん断後にAPPがはじけてしまい、N端のAPPSα(sAPPαとも書ける)とC端のα-CTFが形成されている。この2つの断片が多いほど、APPのせん断が「良性」に近くなり、Aβが発生しにくいと簡単に理解できる。α-CTFの更なるせん断となると、ひとまず置いておく。
アミロイド剪断なら、β-secretase(βカット酵素)によってカットする。AβシーケンスのN端に切り、APPはAPPSβ(sAPPβとも書ける)とC端のβ-CTFに分けられる。先ほど述べたSwe突然変異はAPPのβせん断をより容易にする。これらの「β」の持つものの出現はAβの量の増加を意味する。β-CTFがγカット酵素によって再びカットされた後にAβが正式に登場するから。
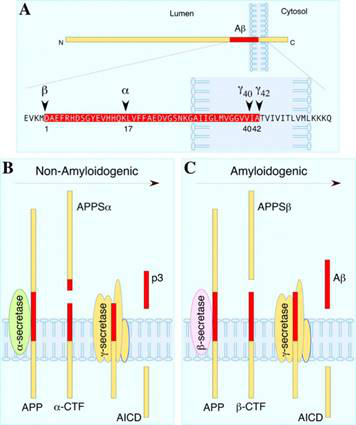
図1. APPせん断のプロセス
ちなみに、γカット酵素はαとβカット酵素と違って、後の2つのカット方法が正確で、位置も固定である。一方、γはα-CTFまたはβ-CTFのせん断に対して不安定である。したがって、Aβは確定なサイズの断片ではなく、1種類の短いペプチド(36-43のアミノ酸も可能である)を指す。要するに、Aβが長いほど集まりやすく、毒性が大きくなる。
2. Aβヒト化突然変異ラット
人類といくつかの霊長類動物のAβ断片だけが凝集しやすいので、Aβ断片をヒト化しないと、ネズミ目に多くの突然変異を導入しても、Aβ凝集物が発生しにくく、モデル構築効果に影響を与えてしまう。そのため、ラットモデルを構築するにはAβ断片をヒト化する必要がある。次に、Aβヒト化したラットを突然変異させる。次に3つのAβヒト化突然変異ラットを紹介する。
Luciano D’Adamio研究室では3種類のAD遺伝子改変ラットを発表した。これらのラットは遺伝子ノックイン(Knock-In)を選択した。なぜ従来の遺伝子組換え(Transgenic,Tg)ではなく、KIを選択するかと質問するかもしれない。従来の遺伝子組換え方式の研究でAD治療薬物(Aβ仮説に基づく)が開発されていなかったためである。一方、Tgに比べて、KIは以下のように3つの利点がある:
1. Tgに比べて、KIはランダムに挿入されておらず、ラット自体の遺伝子への影響は最小限に抑えられている。
2. 多コピーがなく、人類の病気の病理的特徴にもっと近い。
3.ラット遺伝子の元のシーケンスのほとんどを保留し、ラットの既存コントロール元は依然として役割を果たすることができる。
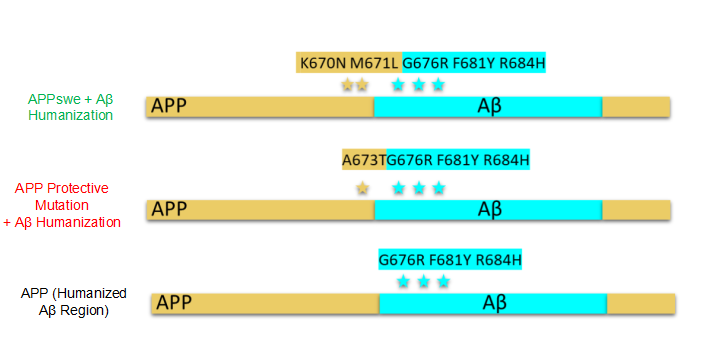
図2. 三種類のAPP遺伝子改変ラット
筆者はAβヒト化したラットに対して2種類の突然変異を行った。一つはありふれたSwe突然変異、つまりK670N/M671L突然変異である。この突然変異は家族性ADの発生をもたらし、患者に認知障害を与えた。もう一つはA673T突然変異である。この突然変異はADの認知障害に一定の保護の役割がある。
Luciano D’Adamio研究室ではPS1突然変異ノックインラットを作成し、同時にヒト化されたAβシーケンスとL435Fのホモ突然変異がある。
![]()
図3. PS1のL435F突然変異部位
Aβヒト化+A673T突然変異ラット
まず、以下の図の各ラットの名前を説明する。w/wは野生型で、δ7/δ7は7つの塩基断片を削除したAPPで、h/hはヒト化Aβホモで、s/sはヒト化に基づいたswe突然変異ホモで、p/pはヒト化に基づいたA673T突然変異ホモである。21日間に、3種類のヒト化ラットのAPPの発現は変わっていない。
p/pのsAPPαレベルは明らかにh/hより高く、sAPPβはh/hより低い。これはラットのA673T突然変異がβカット酵素のせん断を抑制していることを説明している。同時に、β−CTFもそれに応じて減少した。これはA673T突然変異がこのラットの21日間の時にAPPのせん断面で一定の保護役割を持ち、つまりβカット酵素を抑制してAPPをせん断することを説明している。
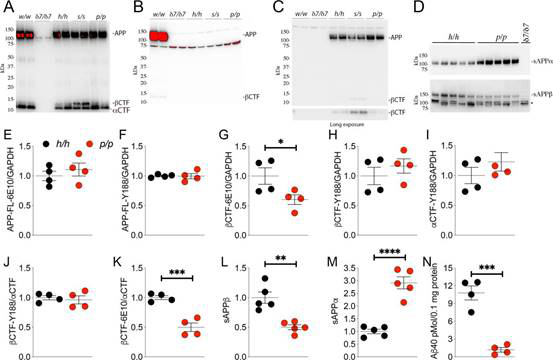
図4. A673T突然変異がβカット酵素のせん断作用を抑制する【2】
βカット酵素の機能を抑制する以外、理論的に、β-CTFの減少により、Aβの発現も抑制されるはずである。予想したとおり、28日間のラットの各種Aβは著しく低下した。めぼしいのは、この研究もラットに対してオスメスの違いが行い、結果はほぼ同じである。WBによるCTFの検測結果もN端生成物と同様である。これはA673T突然変異がAβの産生を抑制することを説明している。興味深いことに、Aβ38の検測で、このAβはメスのみに存在する。
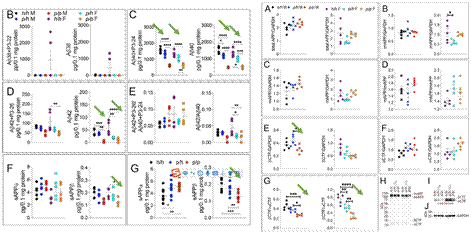
図5. A673T突然変異がAβの産生を抑制する【2】
Aβヒト化+swe突然変異ラット
逆に、APPsweラットであれば、明らかなβせん断とAβ発現の増加がある。同時に、βせん断の増加により、成熟したAPPの含有量も減少した。このため、このラットモデルはAPPのせん断においてADを効果的にシミュレーションした。
要するに、保護的なA673T突然変異はβせん断を減少し、swe突然変異はβせん断を増加する。前者は最後にAβを低下し、後者はAβを増加する。
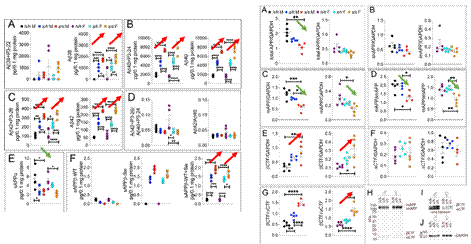
図6. swe突然変異がAβを増加する【2】
Aβヒト化+PS1突然変異ラット
図1のγカット酵素は4つの蛋白質から構成されているが、その中で最も重要な構成要素はプレセニリン1(Presenlin1,PS1)である。Luciano D’Adamio実験室はPS1突然変異ノックインラットを作成した。
このラットはL435Fの突然変異を発現できる。これはマウスと異なる。もしマウスがこのPS1のホモ突然変異を含んでいれば死んでしまい、その後の実験ができない。ホモL435Fラットは27日間の時の体重低下と1月齢時の40%の死亡率を引き起こすことがあるが、一ヶ月を超えたラットは2月齢まで生きても問題ない。そのため、L435Fホモラットは実験を行われる。また、438部位の突然変異は同義突然変異であり、アミノ酸配列に影響がない。
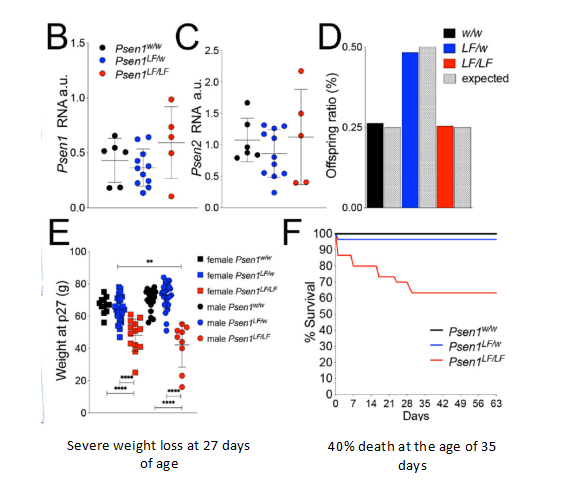
図7. PS1のホモ突然変異がラットの体重と生存率を減少する【1】
さらに、ラットは産後4日間にAβ38、Aβ40とAβ42が低下したが、Aβ43が増加した。同時に、高分子Aβと低分子Aβの比も突然変異の影響で増大するので、この結果はPS1-L435 F KIがγカット酵素の全体せん断能力を低下するが、Aβ43のせん断能力を向上すると説明している。Aβの毒性はAβの豊度と関係しているだけでなく、異なるAβとの比も関係している。この結論がAβの毒性にどのような影響を与えるか、更なる研究が必要である。一方、使用しているラットは4日間生まれたばかりなので、年齢の増加とともにγカット酵素の機能が変化するかは不明である。
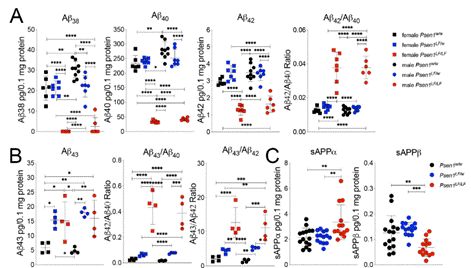
図8. PS1のL435Fの突然変異がAβ43を増加する【1】
以上のように、実はAD病理特徴を持つラットKIモデルを持っている。そして遺伝子技術でラットでの応用が益々成熟していくにつれて、人類のもっと多くの突然変異をラットに導入して、人のAD病理表現型を模擬できる。現在、ADラットモデルの数量は比較的に少ないが、AD研究や神経生物学研究におけるラットの優性を改めて重視しなければならない。
しかし、残念なことに、前述のADラットモデルは病理実験でまだ年齢が小さい(最大の実験年齢も28日間で、最小4日間である)ため、多くの分子レベルの変化に時間がかかる。また、神経生物学において最も重要な行動表現型も一定の年齢後に測定する必要があり、これはモデルの優劣の鍵となる。そのため、今後の行動研究でよい結果を得ることを期待しております。
ラットはマウスより賢いので、行動実験における分解能はさらに向上し、既存の分子と病理レベルの変化は行動に反映しやすい。
また、異なる遺伝子改変モデルが構築されている場合、これらのラットを交配すれば、Tgマウスのような複数の遺伝子ラットも得られ、より明らかな病理表現型を得ることができる。このため、ラットモデルの構築、開発と最適化はADの研究、特に薬物開発研究に強力な武器を提供する。
参考文献:
1.Tambini MD, D'Adamio L. Knock-in rats with homozygous PSEN1L435F Alzheimer mutation are viable and show selective γ-secretase activity loss causing low Aβ40/42 and high Aβ43. J Biol Chem. 2020;295(21):7442-7451. DOI: 10.1074/jbc.RA120.012542
2.Tambini M D , Norris K A , D'Adamio L . Opposite changes in APP processing and human Aβ levels in rats carrying either a protective or a pathogenic APP mutation[J]. eLife Sciences, 2020;9:e52612 DOI: 10.7554/eLife.52612
最新の販売促進キャンペーン:
- Cyagenノックアウトマウスライブラリ:16000種以上のKO/cKO系統マウスを所有、早くて2週間で納品
- 期間限定の特別価格でACE2マウスモデルを提供致します!
- ノックアウトラットモデル:KO、cKO、KI、点突然変異とPiggyBac遺伝子組換え、今なら200万円!
サイヤジェン株式会社について
サイヤジェン株式会社は15年間の発展を経て、全世界の数万人の科学研究者にサービスを提供しており、製品と技術は直接にCNS (Cell、Nature、Science)の定期刊を含む5,200余りの学術論文に応用されています。弊社の「ノックアウトマウスライブラリ」は低価格だけでなく、遺伝子名称を入力すれば、ワンクリックで注文まで操作できます。 ノックアウトマウス、ノックインマウス、コンディショナルノックアウトマウス、トランスジェニックマウス、GFPマウス、免疫不全マウス、無菌マウスなどのカスタマイズサービスを提供する以外、専門的な手術疾患モデルチームがあり、多種の複雑な小動物手術疾患モデルも提供できます。
